
‡These authors share the first authorship.
§These authors share the last authorship.
Academic Editor: Graham Pawelec
Background: Glioblastoma multiforme (GBM) is the most common and malignant primary brain tumor in adults. Novel treatments are needed to counteract the molecular mechanisms of GBM growth and drug resistance. The chaperone system (CS) members are typically cytoprotective but some, termed Hsp, can become pathogenic and participate in carcinogenesis, along with the vascular endothelial growth factor (VEGF), and we investigated them in GBM biopsies and derived cell lines. The objectives were to identify diagnostic-prognostic biomarkers and gather information for developing chaperonotherapy. Methods: Cell lines from GBMs were established, characterized (morphology, growth characteristics, and specific markers), and stored. Chaperones and angiogenic factors [Hsp10, Hsp27, Hsp60, Hsp70, Hsp90, FLT-1 (VEGFR-1), FLK1 (KDR, VEGFR-2), and FLT-4 (VEGFR-3)] were observed in cells by immunofluorescence while the chaperones were measured in tumor tissue by immunohistochemistry. Results: Four cell lines were derived from four different GBMs; the cells were spindle shaped or polygonal and grew at high rates as adherent monolayers or clusters without evidence of contact inhibition. The astrocyte-specific glial fibrillary acidic protein (GFAP); and the neuronal NSE, malignancy VIM, and proliferation PCNA, markers were determined. The cells expressed GFAP but no NSE, indicating that they were primary glioblastoma cell lines, with high levels of Hsp10, Hsp27, Hsp60, Hsp90, and Flk1; and low levels of Hsp70, Flt1, and Flt4. Conclusions: Four cell lines were established derived from four out of ten GBM tumors studied. The cell lines showed intense positivity for chaperones studied and factors connected to malignancy and the tumors showed increased levels of chaperones, making them potential diagnostic-prognostic biomarkers and targets for anti-cancer compounds.
Glioblastoma multiforme (GBM) develops from astrocytes and is the most aggressive primary cancer in human brain. GBM rarely metastasizes, it induces death through invasion of normal brain tissue and by resisting therapies. The 2016 World Health Organization (WHO) Classification of CNS Tumors combines histological and molecular parameters to define various types of gliomas [1]. Diffuse glioblastomas are separated into two groups, with and without mutation of the isocitrate dehydrogenase (IDH) gene, namely, WHO grade IV with and without IDH mutation [2]. These two groups of gliomas have different histopathological features, modes of progression, and prognoses [3]. IDH mutation frequently occurs in secondary glioblastomas, which grow slowly and have a favorable prognosis. The wild-type IDH occurs in older patients in the absence of any lower-grade precursor and have a worse prognosis [4]. Prognosis and clinical progression also differ significantly within the groups, indicating that there are other factors still to be identified that distinguish tumor subgroups. In addition, the location and size of glial tumors determine the clinical presentation, requiring personalized monitoring and treatment strategies.
The distinctive feature of GMB is the recurrence and short survival time after aggressive therapies. This is due to the “angiogenic switch”, consisting of the formation of a dense network of vessels tortuous and hyperpermeable, with production of pro-angiogenic factors, resulting in the uncontrolled proliferation, infiltration, and progression of the tumor. Angiogenesis is driven by mechanisms that lead to activation of the vascular endothelial growth factor (VEGF), one of the most important regulators of tumor growth [5]. The critical role of VEGF in angiogenesis makes it an appealing target for therapeutics. Preclinical data and early clinical trials with agents targeting VEGF prolonged patient survival; however, acquired anti-medication resistance and incomplete VEGF blockade made clear that other more efficacious therapies for GBM would have to be developed [6, 7].
New avenues for research and for developing novel tools for prognostication and
treatment of GBM have been opened by progression in the knowledge of the
chaperone system (CS) and its functions as a physiological entity in health and
disease, including tumors. The CS of an organism, e.g., a human, is composed of
molecular chaperones, chaperone co-factors, co-chaperones, and chaperone
receptors and interactors [8]. Molecular chaperones, the chief members of the CS,
are proteins classified according to molecular weight into groups, encompassing
the following ranges (in kDa):
The data briefly discussed above and many others in the literature support the notion that the CS system is involved in carcinogenesis, both against and for cancer development. Although the CS system components, particularly the molecular chaperones, are typically cytoprotective, if abnormal they can cause diseases, the chaperonopathies [26]. Therefore, it is pertinent to ask about the role of chaperones in GBM as possible biomarkers useful for differential diagnosis, prognostication, and patient follow up. Furthermore, it is highly likely that by revealing the quantitative and distribution patterns of chaperones in GBM in relation to clinical manifestations, response to treatment, and other pathological parameters, will provide clues for developing efficacious treatments targeting the chaperones, namely chaperonotherapy [27, 28]. Consequently, we have been studying the CS in various cancers and, in the research reported here, we conducted an immunohistochemical evaluation in GBM tissue samples of the chaperones Hsp10, Hsp27, Hsp60, Hsp70, and Hsp90. In parallel, we also studied other molecules pertinent to GBM biology, such as Flt1 (VEGFR-1), Flk1 (KDR, VEGFR-2), and Flt4 (VEGFR-3) in primary glioblastoma cell lines derived from patient biopsies. The immediate objective of this project is to reveal features useful for tumor identification and differential diagnosis, while the long-term goal is to accumulate information that could serve as a platform for the development of therapeutic means targeting the pathogenic chaperones.
The patients enrolled in this study underwent surgery for supratentorial, hemispheric GBM at the Neurosurgery Department, Policlinico P. Giaccone, University of Palermo. In all cases, total tumor removal was done-namely, a resection of the entire contrast-enhancing lesion confirmed by MRI. All patients gave their written informed consent to study participation. This study was approved by the Ethics Committee of University Hospital AUOP Paolo Giaccone of Palermo (number 11/2018) in accordance with current legislation and the ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments. The samples were collected between 2019 and 2021; tumor diagnosis was performed according to the fourth edition of the WHO classification of tumors of the central nervous system. In our study, 10 patients with glioblastoma were enrolled at the Policlinico P. Giaccone and tissue biopsy samples were obtained from nine of them. Cell lines were established from 4 of the 9 samples. The mean age of the patients was 57.75 years (ranging from 53 to 64) and their clinical characteristics such as anatomical site, isocitrate dehydrogenase (NADP(+)) 1 IDH1 wild type (WT) or mutant, Ki67, and synaptophysin are summarized in Table 1. IDH mutations, Ki67, p53 and synaptophysin were determined in all glioblastoma cases from the Institute of Pathological Anatomy. In addition, 10 non-pathological samples of cortex from age-matched subjects, were used for comparative analysis, obtained from U.O.S. Forensic Medicine, Policlinico P. Giaccone.
| Patients | Sex | Anatomical site | IDH1 |
Ki67 percentage | Synaptophysin |
| 1 | M | Left frontal lobe | – | 67.17 | Negative |
| 2 | F | Left parieto-occipital region | + | 67.25 | |
| 3 | F | Right frontal lobe | NOS | 67.25 | |
| 4 | F | Right frontal lobe | NOS | 67.40 | |
| 5 | M | Right frontal lobe insular region | +/– | 67.10 | |
| 6 | F | Left frontal lobe | – | 67.40 | |
| 7 | M | Left frontal lobe | – | 67.40 | |
| 8 | M | Left frontal lobe | – | 60.40 | |
| 9 | F | Right temporal lobe | + | 60.20 | |
| 10 | M | Right temporal lobe | + | 67.20 | |
Immunohistochemical staining was performed using formalin-fixed
paraffin-embedded blocks. Briefly, 4-
| Method | Antigen | Antibody | Clone | Supplier | Dilution |
| IHC/IF |
Hsp10 | Mouse monoclonal | clone D-8 | Santa Cruz Biotechnology, Dallas, TX, USA | IHC: 1:100 IF: 1:50 |
| IHC/IF | Hsp27 | Goat polyclonal | clone F- 4 | Santa Cruz Biotechnology | IHC: 1:200 IF: 1:50 |
| IHC/IF | Hsp60 | Rabbit polyclonal | H-300 | Santa Cruz Biotechnology | IHC: 1:300 IF: 1:50 |
| IHC/IF | Hsp70 | Mouse monoclonal | 5A5 | Abacam, Cambridge, UK | IHC: 1:100 IF: 1:50 |
| IHC/IF | Hsp90 | Mouse monoclonal | ab13492 | Abcam | IHC: 1:200 IF: 1:50 |
| IF | GFAP | Mouse monoclonal | 2E1 | Santa Cruz Biotechnology | 1:50 |
| IF | NSE | Mouse monoclonal | MRQ-55 | Ventana Medical System, Tucson, Arizona, USA | 1:50 |
| IF | VIM | Mouse monoclonal | V9 | Biocare, Dallas, USA | 1:50 |
| IF | Mouse monoclonal | TUB 2.1 | Sigma-Aldrich, St Louis, MO, USA | 1:50 | |
| IF | ALDH1 | Mouse monolconal | H-8 | Santa Cruz Biotechnology | 1:50 |
| IF | PCNA | Mouse monoclonal | F-2 | Santa Cruz Biotechnology | 1:50 |
| IF | FLT-1 | Rabbit polyclonal | C-17 | Santa Cruz Biotechnology | 1:50 |
| IF | FLK-1 | Mous emonoclonal | A-3 | Santa Cruz Biotechnology | 1:50 |
| IF | FLT-4 | Rabbit polyclonal | C-20 | Santa Cruz Biotechnology | 1:50 |
Tumor tissue was collected during resection and transferred cooled to the
laboratory. Under sterile conditions, it was rinsed in PBS and, subsequently,
obvious vessels, coagulated blood, and necrotic tissue were removed. Each tumor
specimen was dissociated into small pieces, using scalpels, and dissociated
further enzymatically by incubation with Collagenase Type II (250 U/mL, Sigma
Aldrich, St. Louis, MO, USA), 0.25% of trypsin (Thermo Fisher, Waltham, MA, USA)
in PBS at 37
We used the cell line G166, isolated from malignant glioma (ISENET Biobanking
Cell Lines) that show stem cell properties and with the capability of initiating
high grade gliomas after xenograft. Cells were grown in serum-free complete
medium (Euromed-N, EuroClone; code ECM0883LD), with supplements: 2% B27 (GIBCO;
code 17504-044), 1% N2 (GIBCO; code 17502-048), 20 ng/mL epidermal growth factor
(EGF, Peprotech; code 100-15) added fresh; 20 ng/mL basic fibroblast growth
factor (FGF2, Peprotech; code 100-18B) fresh added; on the laminin coating at 1
Five thousand cells per well were plated in chamber slides and allowed to attach
and proliferate for 24 or 48 h. After fixation (ice-cold methanol
for 30 min, Carl Roth, Karlsruhe, Germany) the cells were stained with Hematoxylin-Eosin (H&E), using stain in hematoxylin solution (Papanicolaou
Harris Hematoxylin, Bio-Optica: code: 05-12011) for 1 h at 22
For immunofluorescence, the cells were placed in eight-well chamber slides,
cultured for 24 h and fixed with ice cold methanol for 30 min. The fixed cells
were washed with PBS pH 7.4 and then incubated with unmasking solution (trisodium
citrate 10 mM, 0.05% Tween 20, pH 6) for 10 min at 22
Statistical analyses were carried out using the GraphPad Prism 4.0 package
(GraphPad Inc., San Diego, CA, USA). Comparisons of immunohistochemical
evaluations were made by using Student’s t-test. All data are presented
as the mean
Hsp10 immunopositivity was cytoplasmic in glioblastoma cells with an average
percentage of positive cells of 61.6
 Fig. 1.
Fig. 1.Immunohistochemical demonstration of molecular
chaperones in GBM tissue. Representative images for Hsp10, Hsp27, Hsp60, Hsp70
and Hsp90 in glioblastoma multiforme (GBM) and control (CTR) tissues. The
staining pattern for the Hsp10, Hsp27, Hsp60, and Hsp90 proteins was cytoplasmic,
with strong intensity in GBM tissue. Low immunopositivity of Hsp70 in the
cytoplasm in GBM tissue like that of CTR. Magnification 400
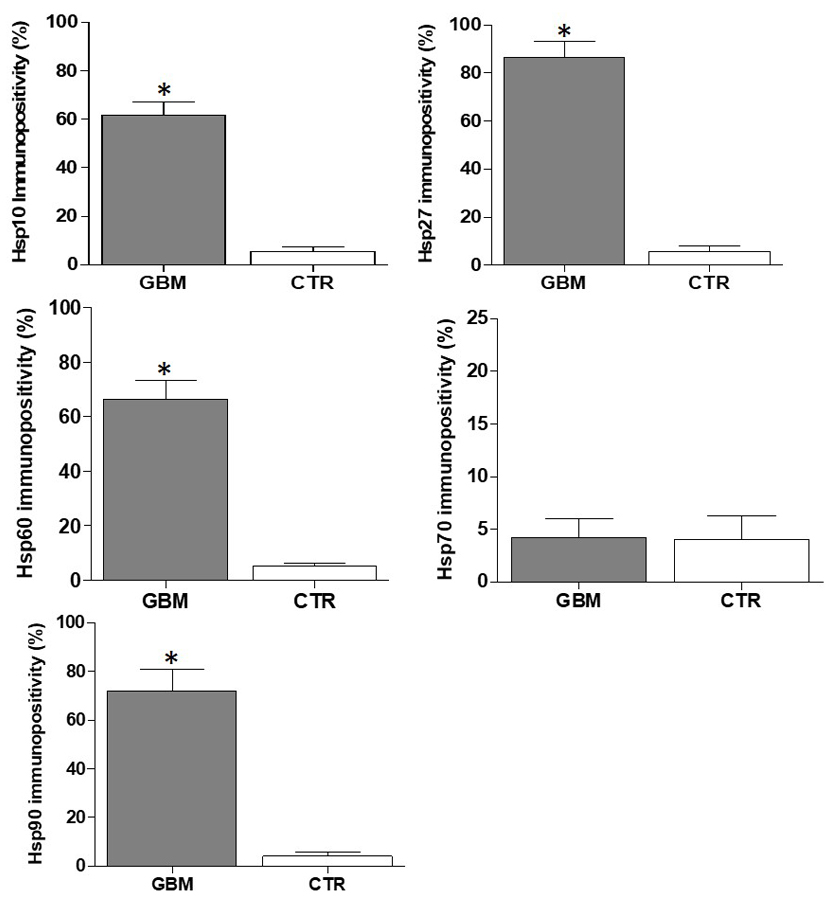 Fig. 2.
Fig. 2.The chaperones tested except for Hsp70 were augmented
in GBM by comparison with the control tissue. The histograms show the percentage
of immunopositivity for Hsp10, Hsp27, Hsp60, Hsp70, and Hsp90 in GBM and CTR
tissues. Data are presented as the mean
The tumor-tissue sample from one of the 10 patients did not pass the short-term primary stage of culture. Of the other nine glioblastoma-tissue samples, five did not attach after disaggregation and did not grow. The other four samples, however, did grow from the start and yielded self-replicating passageable cell lines. This diversity of reaction to explantation probably reflects the area of the tumor from which the sample originates. If the sample is taken from a necrotic area, there are no cells, or extremely few that can grow in the cultures. If the tumor sample originates in a transitional area between necrosis and proliferation, cultivation is possible, but often large amounts of cell debris are present in the culture causing reducing cell viability still more. The presence of erythrocytes in the culture of samples with a high blood supply also leads to cell death. The four cell lines described here were designated GBM1, GBM2, GBM3, and GBM4. The time from explantation (p0) of the tumor samples to the first sub-cultivation step (p1) differed between the primary cell lines: one week for GBM2 and GBM3, and two and three weeks for GBM1 and GBM, respectively.
The new cell lines showed two types of growth: (a) monolayer with the cells growing side by side; and (b) focal with the cells growing in clusters or islands connected by cell protrusions. The commercial cell line G166 grew as a monolayer. In slides stained with Hematoxylin-Eosin, the primary cell lines showed three different cellular shapes: polygonal (Fig. 3A,B), spindle or glia-like (Fig. 3C–F), and amorphous with cellular protrusions (Fig. 3G,H). Multinucleated giant cells were also present at low frequency. The G166 cell line showed a glia-like morphology (Fig. 3I,L). Aliquots of the cell lines have been stored frozen and they recovered and grew well after thawing.
 Fig. 3.
Fig. 3.Morphology of our four GBM cell lines along with that of the
commercially available G166 line revealed by Haematoxylin-Eosin staining. Cell
lines images: GBM1 (A: GBM1 magnification 200
To establish criteria for the definition of primary glioblastoma cell lines and
to ascertain whether the glioblastoma-derived primary cell lines have
similarities to glioblastoma tissue, we resorted to immunofluorescence staining
for markers used in glioma diagnosis with confocal microscopy, comparing our cell
lines with the G166 line. The primary cell lines were tested to determine the
presence of GFAP, which is characteristic of astrocytes or glial cells [30];
Vimentin, a neuroepithelial precursor marker [31];
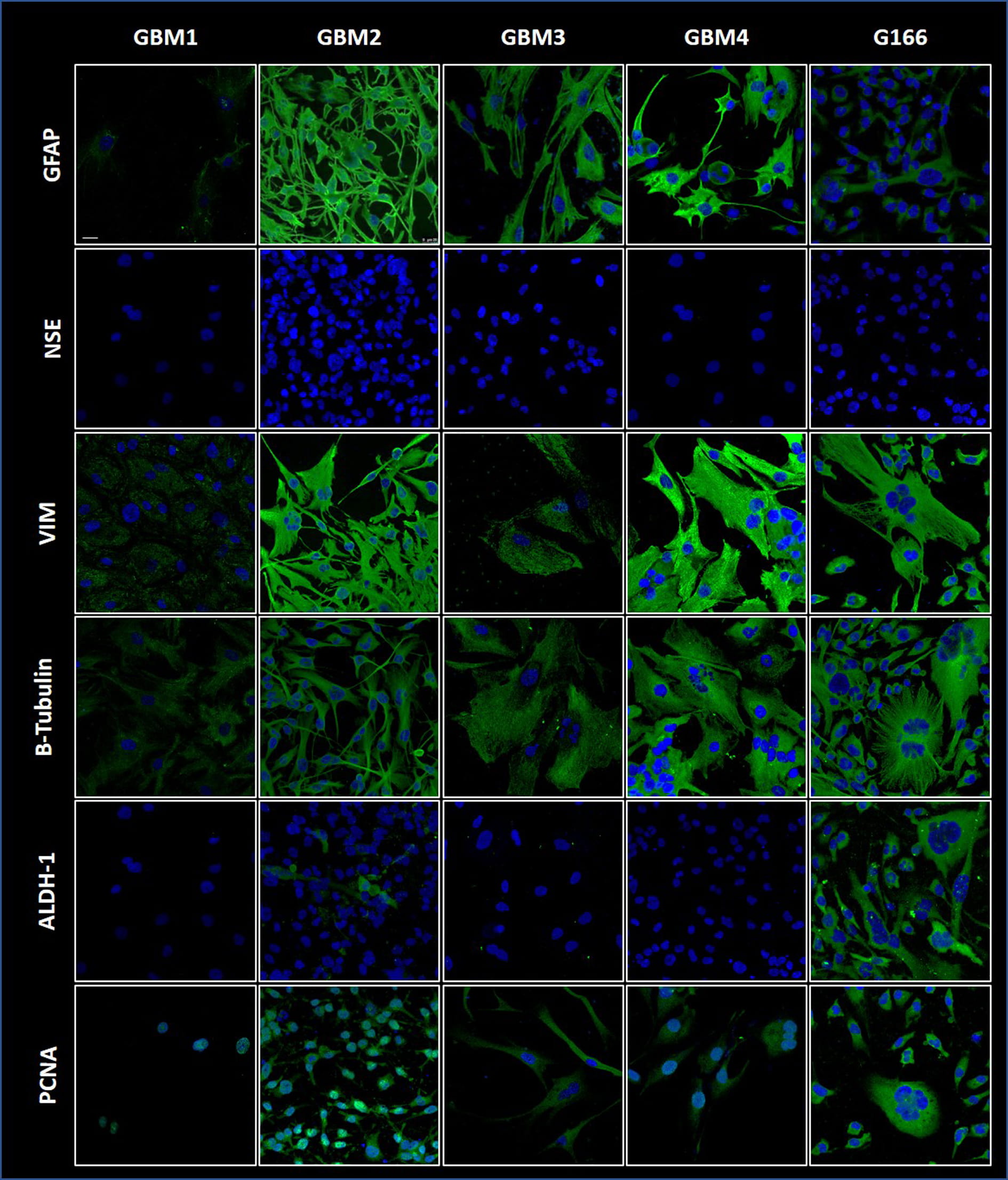 Fig. 4.
Fig. 4.GBM markers detected by immunofluorescence in the four cell
lines along with the G166 line. GFAP, VIM,
The immunopositivity for the molecular chaperone Hsp10 was observed in the cytoplasm of the primary cell lines (Fig. 5) and the cell line that showed the most intense reaction was GBM3, which derived from the youngest patient in the group studied. Hsp27 immunopositivity was intense in GBM2 (this line derived from the GBM patient with the shortest survival after surgery) and occurred also in GBM1, GBM3, GBM4 (derived from GBM patients with longer survival period after surgery) (Fig. 5). Hsp27 staining in the tumor tissue was not significantly different among the GBM patients. The Hsp60 reaction showed a granular and diffuse cytoplasmic positivity in our four GBM cell lines and in the G116 line (Fig. 5). The immunopositivity of Hsp70 was very low or absent in our four cell lines but it was present at very low levels in the G166 cell line (Fig. 5). These results obtained with immunofluorescence are in line with the immunohistochemical data (Figs. 1,2). Our four cell lines and G166 were positive for Hsp90 (Fig. 5). G166 cell line showed cytoplasmic positivity for all Hsps, including Hsp70 (Fig. 5).
 Fig. 5.
Fig. 5.Demonstration by immunofluorescence of CS components in the GBM
cell lines. The chaperones Hsp10, Hsp27, Hsp60, and Hsp90 were present at
diverse levels in our four cell lines, and in the G166 line. Hsp70 was at very
low levels in the four cell lines, comparable to that in C166 or lower. All the
chaperones were present in G166. Specific primary antibodies for the chaperones
and secondary antibody conjugated with FITC (green fluorescence in the figure) or
with TRITC (red fluorescence in the figure) were used, and the cell nuclei (blue)
were stained with DAPI. Magnification 400
Illustrative images of the immunofluorescence mapping of the VEGFR1 (Flt1), VEGR2 (Flk1), and VEGFR3 (Flt4) are shown in Fig. 6. The four GBM lines and the G166 cell line were positive for VEGFR-1 (Flt1) and for VEGFR-2 (Flk1) in the cytoplasm. VEGFR-3 (Flt4) occurred in the G166 cell line and in our cell lines GBM2, 3, and 4 but was undetectable in GBM1.
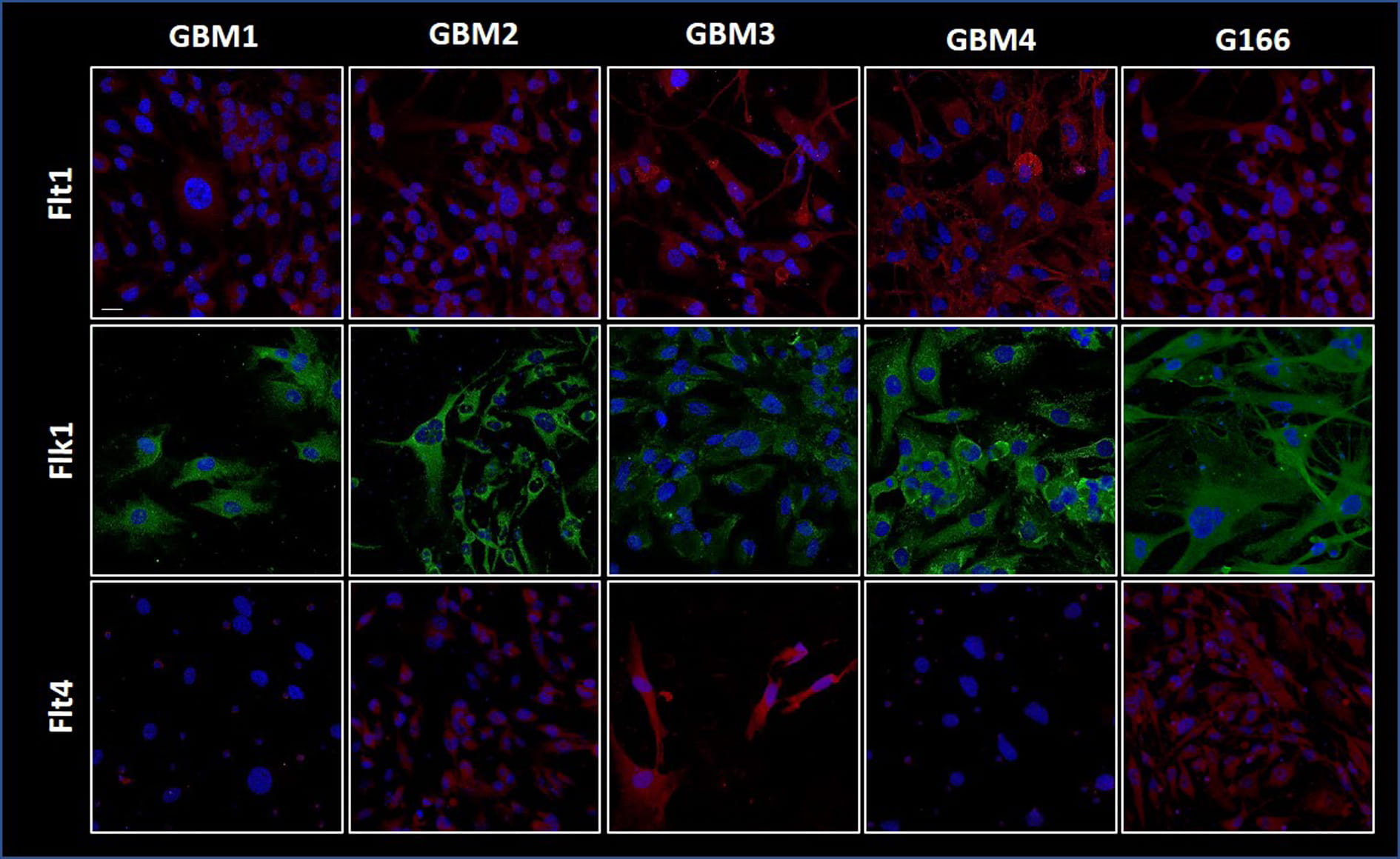 Fig. 6.
Fig. 6.Angiogenesis factors in the GBM cell lines.
Immunofluorescence images for Flt1 (VEGFR-1), Flk1 (VEGFR-2), and Flt4 (VEGFR-3)
showing their presence in the GBM and G166 cell lines. Specific primary
antibodies for the receptors and secondary antibody conjugated with FITC (green
fluorescence in the figure) or with TRITC (red fluorescence in the figure) were
used, and the cell nuclei (blue) were stained with DAPI. Magnification
400
GBMs show a remarkable molecular heterogeneity that defeats the effectiveness of therapeutic approaches that are not patient specific. Understanding the cellular and molecular mechanisms of malignant growth in GBM will be instrumental for identifying new targets for treatment that could guide the development of personalized therapies. In this study, we focused on molecular chaperones. These are the chief members of the CS and are known to play key roles in carcinogenesis, both anti- and pro-tumoral roles [11, 14, 29, 35, 36, 37, 38, 39]. They are also promising candidates as biomarkers for differential diagnosis, prognostication, and patient follow up. We evaluated by immunohistochemistry and immunofluorescence the presence and subcellular localization of the chaperones Hsp10, Hsp27, Hsp60, Hsp70, and Hsp90 in biopsies of ten GBMs. Our experiments showed a high Hsp27 immunopositivity in the cytosol of all GBMs, by comparison with normal tissue. Hsp27 protein levels were higher in biopsies from GBM cases with short survival, a trend found also in other tumors, suggesting that elevated levels of Hsp27 may predict poor prognosis [40]. The immunopositivity for Hsp10 and Hsp60 was also elevated in the GBM tissue in contrast with the normal controls, with Hsp60 present in the cytoplasm, as shown for other types of solid tumors, in which the levels of this chaperonin correlated with tumor progression [15, 41, 42]. Hsp60 and Hsp10 are typically mitochondrial molecules, therefore, our finding them in the cytoplasm of GBM cells supports the notion that the chaperonins augment in the cytosol of tumor cells and are markers of malignancy. The chaperone Hsp70 have been found increased or decreased in human cancers by comparison with healthy control tissues [43, 44, 45]. An inverse correlation between Hsp70 levels and prognosis has been observed in some cancers [35, 46]. In contrast to these findings, we did not detect a clear positivity reaction for Hsp70 in the GBM tissues we describe here, although we found low but still detectable levels of this chaperone in the commercially available stem line G166. This probably indicates that in this stem-cell line, Hsp70 is in the stem-cell nuclei, but it is absent in the other cell lines, in which stem cells have not been demonstrated. Hsp90 was detected many years ago in GBM [47, 48]. In our study, we observed a higher Hsp90 immunopositivity in tumor tissue compared to normal brain tissue. All the observations on the chaperones in the tumor and derived cell lines by immunohistochemistry were confirmed by immunofluorescence.
The four cell lines reported here show some morphological heterogeneity, but most cells have a spindle- or glia-like shape and there are also few multinucleated giant cells. The proportion of these cell types remained unchanged during cultivation and successive passages. The morphological heterogeneity of our cell lines reflects that of the original GBM tumors, which is characteristic of them. The cell lines displayed the markers of neuroglia, for instance GFAP and VIM, indicating that the cells derive from transformed glial cells. Available information suggests that the expression of GFAP and VIM is mostly confined to glial-derived tumors, and that VIM can potentially be a useful marker for distinguishing undifferentiated GFAP-negative glial tumors. Other markers we measured in the cell lines are NSE, ALDH1, and PCNA. We observed no reaction for NSE, a low positivity for ALDH1, and positivity for PCNA. The absence of NSE, a neuronal marker, confirms the non-neuronal origin of the cell lines. ALDH1 has been suggested as a novel stem-cell marker in a small series of human glioblastomas [34], but the expression and functions of ALDH1 in the central nervous system (CNS) and its tumors are largely unknown. Our experiments showed that the primary cell lines do not have stem cell characteristics in contrast with G166, which showed immunopositivity for ALDHI. The immunofluorescence results with anti-PCNA antibodies indicate that the cell lines do show the parameters of malignancy that PCNA represent.
We observed positivity for Hsp27 in the cytosol of the cells and a diffuse positivity for Hsp60 and Hsp10 in the cytoplasm of all cell lines. The positive immunofluorescence reaction of Hsp10 and Hsp60 in the cytosol of the cells matched results obtained by immunohistochemistry in the tumor tissues. The fact that these chaperones were elevated from the first passages of cell growth, and remained elevated during cultivation, suggests that they are implicated in the onset and progression of carcinogenic mechanisms. It is possible that the increase in the amounts of Hsp27, Hsp10, and Hsp60 reflects the exaggerated requirements of tumor cells for new proteins correctly folded caused by their accelerated metabolism and growth and replication rates. The prognostic value of Hsp90 levels depends on the tumor type [48]. Hsp90 is often increased and associated with poor prognosis in various tumors, including GBM [49, 50, 51, 52, 53, 54]. We found high levels of Hsp90 in the GBM2 and GMB4 cell lines with a cytoplasmic localization, in all passages. High levels of Hsp90 may indicate poor prognosis in some cases of GBM, considering that this chaperone can inhibit apoptosis [55].
GBMs are also characterized by neo-angiogenesis and invasion, which is a histopathological hallmark of these cancers associated with a worse prognosis [56, 57]. We therefore examined the expression of the VEGFRs belonging to a superfamily of RTK (receptor tyrosine kinase) and considered key mediators of angiogenesis [58]. VEGFRs are augmented in grade IV glioma vasculature and grade IV glioma cells [59]. Previous studies found that VEGF and its corresponding receptors VEGFRs are significant prognostic factors for overall survival in patients with glioma [60]. Our results demonstrated positive VGFR-1 and VGFR-2 reactions with cytosol localization in the cell lines and a lower positive reaction for VEGFR-3 in the tumor biopsies. Previous studies have shown that in normal brain a low or undetectable endothelial expression of VEGFR-2 can be found but in GBM it increases in parallel with tumor grade [61].
In summary, the levels of Hsp10, Hsp27, Hsp60, and Hsp90 were easily detectable in the GBM tumor tissue, even elevated some of them as compared with normal controls. In contrast, the levels of Hsp70 did not differ between the tumors and the normal tissue, being very low in all samples. These patterns of quantity and distribution of chaperones, if determined in many patients might, in the future, help in prognostication in GBM [60, 62, 63].
Our data provide a platform to elaborate a working hypothesis for future research, considering current knowledge about the CS and its roles in health and disease, including cancer. Hsp90 is a chaperone with a range of client proteins, including oncogene products (e.g., p53, AKT, RAF, EGFR, and MEK) involved in growth, survival, and cell-cycle regulatory pathways [64, 65]. Hsp90 is upregulated within the hypoxic niche of human glioblastomas and a high level of Hsp90 is linked to the upregulation of HIF targets and CSC (cancer stem-like cells) marker genes [25] (Fig. 7).
 Fig. 7.
Fig. 7.Cartoon showing molecules and pathways that are relevant to the biology of GBM and could serve as a platform for a working hypothesis to be tested in search of specific targets, for instance chaperones, for anti-cancer compounds. Left panel; GBM cell. Hsp90 is at the center of pathways to maintain protein homeostasis, including interaction with protein degradations machineries that remove misfolded or otherwise defective proteins and generate building blocks, i.e., amino acids, for the new ones required in larger-than-normal amounts by cancer cells. Hsp90 also participates in tumorigenesis via the HIF/VEGF pathway. For example, hypoxia in the tumor mass induces HIF expression and activity, leading to VEGF production and angiogenesis. The chaperonins Hsp60 and Hsp10 are crucial for maintenance of the mitochondrial functional proteome and, thereby, ensure provision of energy to satisfy the needs of the tumor cell. In addition, Hsp70 and Hsp27 upregulate HIF activity increasing VEGF secretion. It is very likely that the CS, including the chaperones just mentioned, is key for chaperoning the protein molecules mediating the various pathways that are so important to satisfy the “greed” of the tumor cell for energy and proteins. The chaperones assist the tumor-cell proteins to mature and acquire a fully functional conformation and help them to translocate to the place in which they are to function. Therefore, components of the CS may be selected as targets for inhibition, namely negative chaperonotherapy, to hinder tumor survival and stop tumor growth and spread. Right panel; vascular endothelium cell. In vascular endothelial cells, Hsp90 interacts with members of the VEGF-regulated signaling pathway, including VEGFR2. A Hsp90/VEGFR2 complex exists in unstimulated endothelial cells [67]. However, binding of Hsp90 to VEGFR2 increases upon stimulation with VEGF, thus stimulating more angiogenesis. VEGF/VEGFR-2 promotes angiogenesis via a paracrine loop between cancer cells and vascular endothelial cells nearby. In addition, VEGFR-2 in vascular endothelial cells support tumor growth directly or through downstream activation of other signaling pathways, including the SAPK2/p38 pathway. The latter, once activated, triggers into action downstream molecules, such as Hsp27, which involves phosphorylation and stimulating actin remodeling and actin-based motility both necessary for angiogenesis. Hsp90 inhibition could alter the endothelial response and diminish or block angiogenesis by stimulating VEGFR2 proteolysis, providing another argument in favor of the use Hsp90 inhibitors as anti-cancer agents in GBM.
Tumors with elevated Hsp90 had increased levels of the HIF target gene VEGF and
high levels of the glioma stem-cells markers CD133 and nestin, supporting the
notion that Hsp90 is activated by the CSC microenvironment [25]. Hsp90 and Hsp70
interact and regulate various transcription factors, signaling molecules, and
kinases that are related to cancer, including NF-
In conclusion, GBM is a tumor that is still little known in many ways, very aggressive and resistant to current therapies. It is necessary to find new therapeutic strategies that can improve treatment and follow-up tools. New avenues for research on GBM have been opened by progression in the knowledge of the chaperone system (CS) and its functions as a physiological entity in health and disease, including tumors. In this study we conducted an immunomorphological evaluation in GBM tissue samples and cell lines of some molecular chaperones and some angiogenic factors to investigate on their role as biomarkers useful for differential diagnosis, prognostication, and patient follow up. Therefore, on the one hand we wanted to provide useful details on the biology of GBM, on the other hand to add information that could be useful for the development of new therapeutic strategies. Finally, considering the current knowledge on CS and its roles in health and neoplastic diseases, we have developed a working hypothesis to be tested in search of specific targets, for instance chaperones, for anti-cancer drug compounds.
GA, CCB, CC and FR designed the research study. GA, LP, RP and AP performed the research. RB and FR analyzed the data. FG, AMF, and AA provided the samples. GA, RB, FR, wrote the original draft of the manuscript. AJLM, ECdeM, FB and FC wrote, revised, and edited the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
This study was ethically approved by the Ethics Committee of University Hospital AUOP Paolo Giaccone of Palermo (number 11/2018) in accordance with current legislation and the ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments.
AJ.LM and EC deM. were partially supported by IMET and IEMEST. This is IMET contribution number 22-101.
This research received no external funding.
The authors declare no conflict of interest.