
†These authors contributed equally.
Academic Editor: Graham Pawelec
Aims: Intrahepatic cholangiocarcinoma (ICC) is a highly malignant and heterogeneous cancer with a poor prognosis. At present, there is no optimal treatment except for surgical resection, and recurrence after resection will lead to death due to multidrug resistance. Changes in the redox signal have been found to be closely related to the growth and drug resistance of tumor cells. Therefore, the purpose of this study was to screen small molecule compounds from the redox library to find a drug for anti-ICC and to explore its downstream mechanism. Material and methods: Tumor clone and sphere formation of ICC cell lines, as well as mouse ICC organoid proliferation assays were utilized to screen the candidate drug in the Redox library. Western blotting, quantitative reverse-transcription polymerase chain reaction (qRT-PCR), as well as cell apoptosis and cell cycle flow cytometry assays were used to explore the mechanism. Results: We found that Hinokitiol was a candidate drug through inhibition of tumor clone and sphere formation, and the expression of cancer stem cell (CSC)-related genes. Furthermore, Hinokitiol significantly inhibited the proliferation of ICC cells by downregulating the ERK and P38 pathways. In addition, the combination of Hinokitiol and Palbociclib showed a significant inhibitory effect on human ICC cells and mouse ICC organoids. Conclusion: Hinokitiol may have the potential to be developed as a clinical therapeutic drug for ICC treatment.
Intrahepatic cholangiocarcinoma (ICC), which originates from intrahepatic bile duct epithelial cells, is the second most common human primary liver cancer after hepatocellular carcinoma (HCC) [1, 2]. Recently, studies have shown an increasing incidence and mortality rate of ICC, especially in Eastern Asia [3]. Although surgical resection is the most effective treatment for early stage ICC, only a few patients are suitable for this therapy [4, 5], as the majority of ICC patients are diagnosed at an advanced stage, lacking the opportunity for resection [6], and recurrence leading to multidrug resistance is the key problem of ICC treatment [7, 8]; therefore, it is essential to develop a chemotherapy regimen to overcome this refractory malignancy.
Redox signaling has been reported to be a feasible target for overcoming multidrug resistance in cancer chemotherapy [9, 10]. Changes in the oxidation-reduction equilibrium state may lead to oxidative stress and abnormal cell signal transduction [11, 12], which in turn regulate redox-sensitive enzymes and transcription factors [13]. It can also directly affect the curative effect of cancer treatment through various mechanisms, such as apoptosis, growth inhibition [14], angiogenesis, and metastasis [15, 16]. For example, higher ROS levels could promote the development of cancer by inducing DNA mutations [17, 18], genome destruction, and abnormal signal transduction of tumor initiation, proliferation, and drug resistance [19]. Detoxifying enzymes and antioxidant proteins in redox-related pathways play a key role in regulating the balance between apoptosis and carcinogenesis [20]. Moreover, the overactivation of nuclear factor-erythroid 2 related factor 2 (Nrf2), a redox-sensitive factor, may be involved in the occurrence of breast cancer and ovarian cancers [21, 22]. Apurinic-apyrimidinic endonuclease 1/redox factor 1 (APE-1/Ref-1) [23], another redox factor, can promote the binding of transcription factors to gene promoters involved in cancer initiation and progression. These redox-related factors may be responsible for the occurrence and development of tumors. Therefore, regulating the redox levels of tumor cells may be of great significance for the development of potential anti-cancer therapies.
The redox library in our study contains a variety of 84 small molecular chemical compounds that inhibit the redox reaction. In this study, we explored whether these small molecules in the redox library can effectively inhibit the clone formation and tumor sphere formation in ICC cells. We screened the redox library and found that #15 Hinokitiol was the candidate molecule that could inhibit the cancer stemness of ICC cells. Using qRT-PCR assays, flow cytometry, and western blotting analysis, we found that some CSC-related markers were downregulated by #15 Hinokitiol treatment through inhibition of the ERK and P38 pathways. Based on the functional role of Hinokitiol in the repression of proliferation and tumor sphere formation of ICC cells, Hinokitiol has the potential to develop into a new anti-cancer drug with good biological safety.
All animal experiments in this study were conducted in accordance with ethical regulations and approved by the Ethics Committee of the Academy of Military Medical Sciences.
Intrahepatic cholangiocarcinoma cell line HuCCT1 cell was provided from Beijing
Beina Chuanglian Biotechnology Institute. RBE and HCCC-9810 cell lines were
obtained by China Infrastructure of Cell Line Resource (CICR). QBC939 cell line
was given by Prof Jiahong Dong (Beijing Tsinghua Chang Gung Hospital, Beijing,
China). ICC cell lines (QBC939 and HuCCT1) were grown in high glucose DMEM medium
supplemented with 10% FBS (Invitrogen), 100
Redox library was established from Center for Excellence in Molecular Cell Science, Chinese Academy of Sciences (Shanghai, China), as shown in Supplementary Table 1. All the small molecular drugs were dissolved in DMSO and the storage concentration was 10 mM.
A total of 1500 ICC cells were seeded in the 24-well plates (Corning, USA) and
cultured in the DMEM medium supplemented with 10% FBS (Invitrogen). After 24 h,
ICC cells were treated with 84 small molecules in the redox library with a
concentration of 10
A total of 3000 ICC cells were seeded in the 6-well ultra-low attachment plates
(Corning, USA) and cultured in the DMEM/F-12 supplemented with B27 (Life
Technologies, USA, 1:50), N2 (Life Technologies, 1:100), 20 ng/mL epidermal
growth factor (EGF), 10 ng/mL bFGF, 100 units/mL penicillin, and 100 ng/mL
streptomycin and seven preliminary screening small molecules with a concentration
of 10
Paraffin-embedded tissue sections were dewaxed and hydrated in alcohol, then,
10% hydrogen peroxide was added for 30 min to remove endogenous peroxidase.
After induction of Antigen epitope retrieval by microwave heating. Tissues were
immunostained with primary antibodies overnight at 4
The cell apoptosis was detected with Annexin V, FITC Apoptosis Detection Kit
(Dojindo, #AD10-10). A total of 2
A total of 1
Total RNAs were extracted from the cells with TRIzol (Invitrogen). The amount of
RNA was quantified spectrophotometrically with a Nano-Drop ND-1000. Reverse
transcription of total RNAs into cDNA was performed with a reverse transcriptase
kit (FSQ-201, TOYOBO, Japan). The relative mRNA expression was determined by real
time PCR using THUNDERBIRD SYBR qPCR Mix (QPS-201, TOYOBO, Japan). Briefly, a 20
The ICC cells were lysed with RIPA buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl,
1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS) supplemented with protease
inhibitors (Calbiochem, San Diego, California), and equal amounts of protein was
separated by SDS-PAGE (10% polyacrylamide), and then transferred the protein to
polyvinylidene difluoride membranes (PVDF, BioRad, Hercules, CA). The membranes
were incubated with primary antibody overnight at 4
C57BL/6 mice of 8~10 week old were purchased from Vitalriver
(Beijing, China) and raised under specific pathogen-free conditions. The plasmids
ddAKT and ddYAP were kindly gifted by Prof Yuji Nishikawa (Asahikawa Medical
University, Asahikawa, Hokkaido, Japan). The plasmid containing Sleeping Beauty
transposon (SB13) was preserved in the laboratory. These three plasmids were
dissolved in 2 mL saline and injected into the mouse via tail vein injection
(5~7 s). After 8~10 weeks, the abdominal bulge
was observed and the mice were euthanized. The tumor tissue (about
3~5 mm
Isolated primary mouse ICC cells were dissolved in the organoid culture medium
(Stemcell, #06030) and mixed with Matrigel (BD, #356234) in precooled EP tube and
and suspended the pellet at 4
We analysed the data using GraphPad Prism software version 8 (GraphPad Software,
San Diego, California, US). Comparisons between two groups were performed with
unpaired t tests. All experiments were conducted at least three times
and data are presented as means
The anti-tumor effect of 84 different small molecules in the redox library with four human ICC cell lines were preliminary screening using clone formation assays (Fig. 1A–D). The representative clone formation of preliminarily screened four ICC cell lines is displayed in Fig. 1E, Supplementary Fig. 1A–C. We statistically analyzed and merged the inhibitory effect on four ICC cell lines treated with 84 different small molecules in the redox library, as shown in Fig. 1F. Eight small molecule inhibitors (#3, #15, #40, #54, #61, #67, #68, and #71) showed a significant decrease in clone formation capacity compared with the control group treated with DMSO. Among them, #54 (n-Octylcaffeate) has not been sold in China, so we will not consider using this drug for further exploration. These preliminary results indicated that these seven small molecule inhibitors could significantly suppress the proliferation of human ICC clones.
 Fig. 1.
Fig. 1.Preliminary screening of drugs from the redox library via clone
formation assays. (A–D) The column graph of HuCCT1 (A), QBC939 (B), RBE (C), and
HCCC-9810 (D), each treated with small molecules in the redox library, compared
with the control group. (E) Photos of the clone formation assay of HuCCT1 cells
treated with various drugs in the redox library. (F) Venn diagram of anticancer
drugs (with over 50% anti-tumor effect) in four ICC cell lines treated with 84
drugs from the redox library. Data was analyzed using unpaired t test.
N.S., no significance, *p
The tumor sphere formation assay is a standard method for evaluating the
proliferation ability of CSCs in vitro. The above seven candidate small
molecular inhibitors were further screened for their effects on tumor sphere
formation in four human ICC cell lines (Fig. 2A–E, Supplementary Fig.
2A–C). The number of tumor spheres treated by these candidate small molecules
was statistically analyzed using a column graph and Venn diagram (Fig. 2F). The
results showed that
 Fig. 2.
Fig. 2.Screening of drugs from the redox library via tumor sphere
formation assays. (A–D) The column graph of the tumor sphere formation of HuCCT1
cells (A), QBC939 cells (B), RBE cells (C), and HCCC-9810 cells (D), each treated
with small molecule compounds from redox library. (E) Photos of the tumor sphere
formation in QBC939 cells treated with small molecules in the redox library;
Scale bar = 250
Drug resistance caused by CSC enrichment has been proven to be the main cause of
chemotherapy failure and cancer recurrence in various cancers [24]. We detected
the altered expression of CSC-related genes in human ICC cells treated with four
candidate drugs at the secondary screening stage using qRT-PCR assays (Fig. 3A,B,C,D). Compared to the control group treated with DMSO, we found that several
CSC-related markers, excluding KLF4 and CD24 in HuCCT1 and
QBC939 cells, were significantly downregulated by #15 Hinokitiol, not by #3
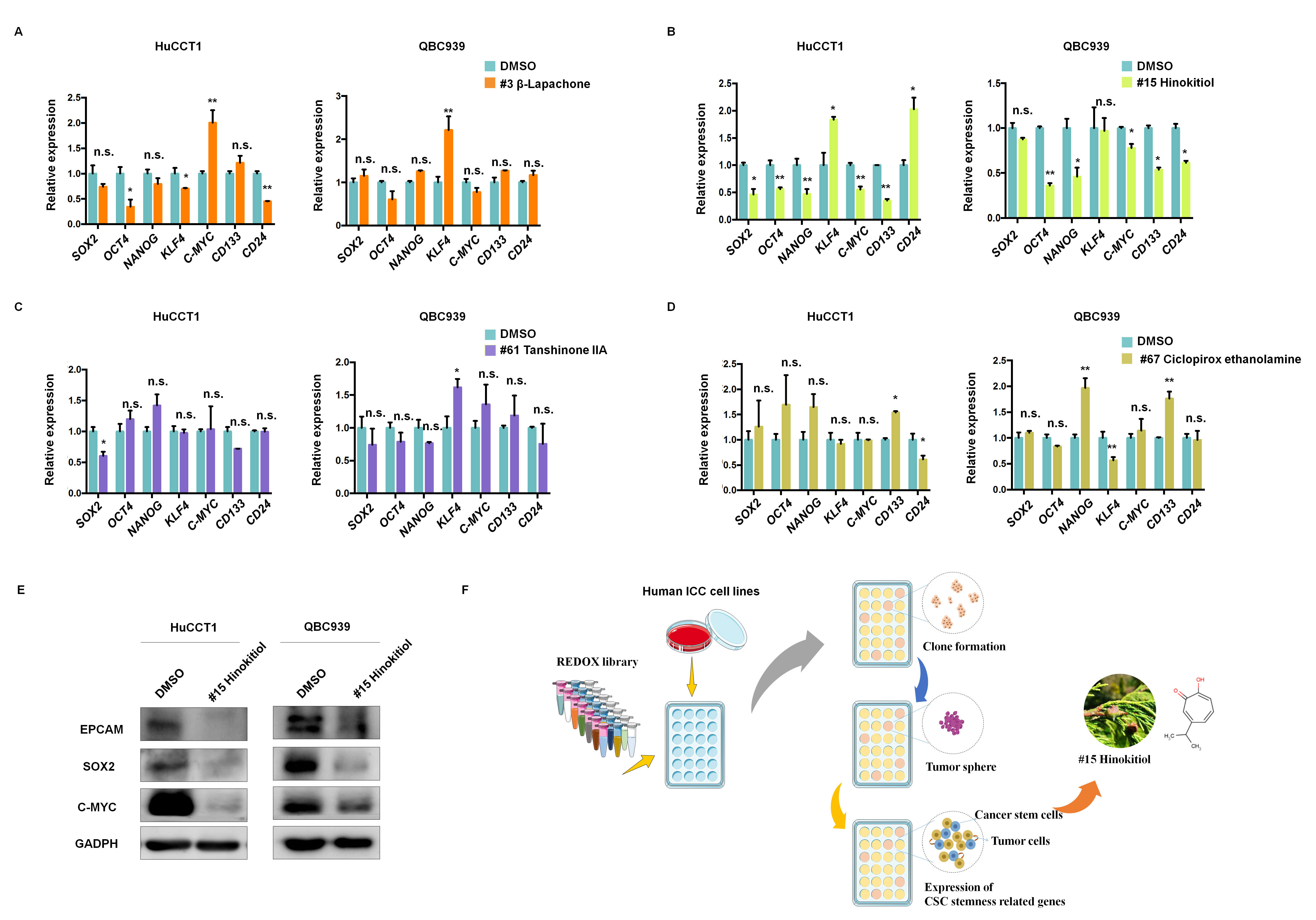 Fig. 3.
Fig. 3.Hinokitiol significantly inhibited some CSC-related markers.
(A–D) Relative expression of CSC-related genes in HuCCT1 and QBC939 cells
treated with
Energy-dependent drug efflux, cell cycle promotion, and apoptosis escape are usually related to the drug resistance in CSCs [24]. To explore the downstream mechanism of ICC cells treated with Hinokitiol, we performed apoptotic and cell cycle analysis of ICC cells after Hinokitiol treatment using flow cytometry. As shown in Fig. 4A,B, there were no significant differences in the early and late apoptotic cell populations between the Hinokitiol-treated group and control cells. The cell cycle analysis also showed that there were no significant differences between the Hinokitiol-treated group and the control group in G1, G2, and S phrase (Fig. 4C,D). These results suggest that the concentration of Hinokitiol used in our experiments can inhibit the proliferation of ICC cells and CSCs (Supplementary Fig. 3A,B), but not through apoptosis induction or cell cycle blockage.
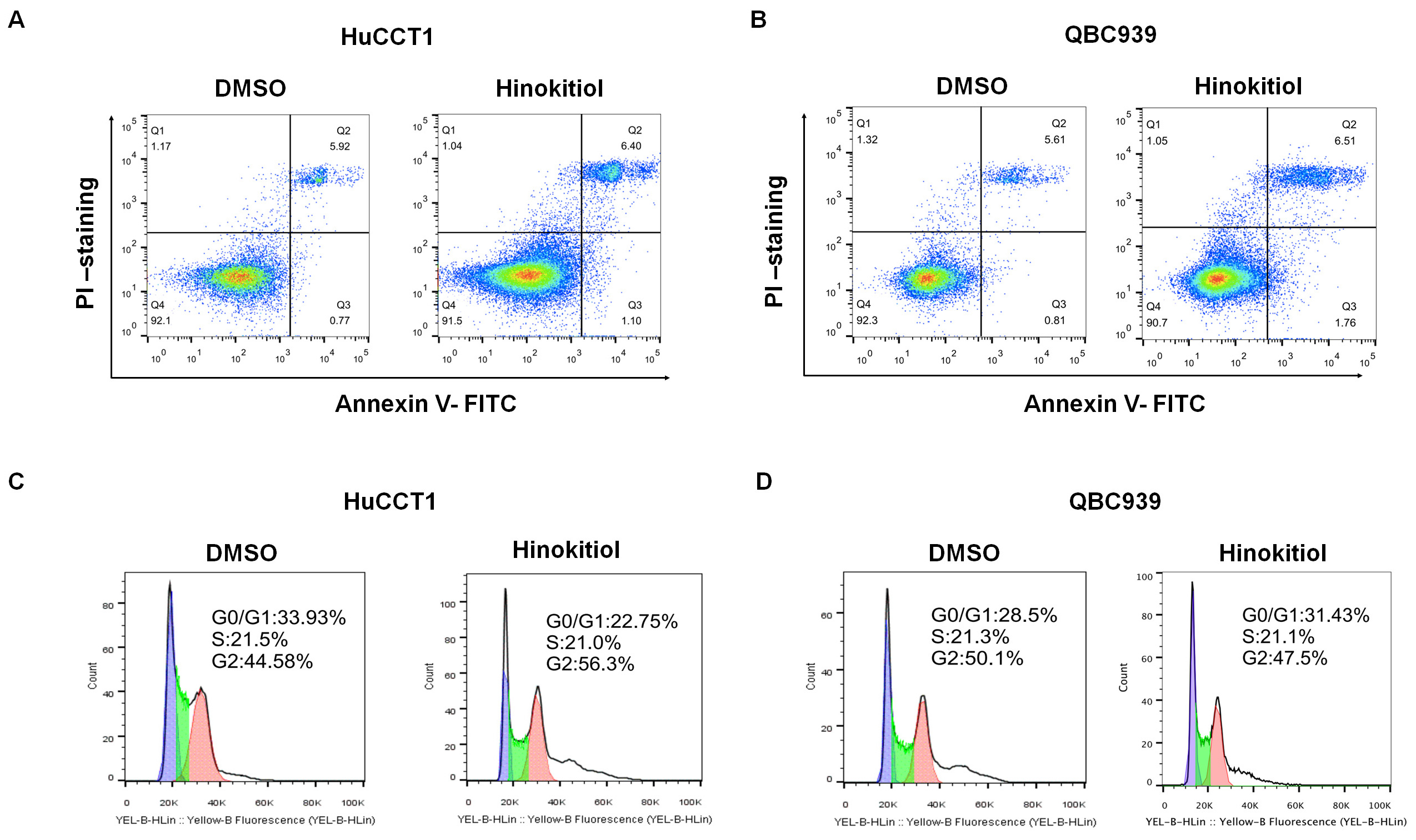 Fig. 4.
Fig. 4.Effects of Hinokitiol on the apoptosis and cell cycle of ICC cells. (A) Apoptosis of HuCCT1 cells treated with DMSO and Hinokitiol. Q1, Q2, Q3, and Q4 population represents the number of living cells, the number of late apoptotic cells, the error within the detection range, the number of early apoptotic cells, respectively. (B) Apoptosis of QBC939 cells treated with DMSO and Hinokitiol, respectively. (C,D) Cell cycle of HuCCT1 and QBC939 cells treated with DMSO and Hinokitiol, respectively.
To further investigate the underlying mechanism of Hinokitiol-induced CSC repression in human ICC cells, we detected the expression of key target genes in various signaling pathways by qRT-PCR assays, including WNT, Hedgehog, NOTCH, mitogen-activated protein kinase (MAPK), and epithelial-mesenchymal transition (EMT) (Fig. 5A,B). As shown in Fig. 5B, the expression of ERK2 and P38, which belong to the MAPK signaling pathway, exhibited a remarkable reduction in HuCCT1 and QBC939 cells in the Hinokitiol-treated group. Moreover, the expression levels of total and phosphorylated ERK and P38 proteins were further verified by western blot analysis (Fig. 5D). Overall, these results indicate that Hinokitiol may reduce the proliferation of human ICC cells by decreasing the expression of ERK, p-ERK, P38 and p-P38.
 Fig. 5.
Fig. 5.Hinokitiol down-regulated the expression of ERK and P38 pathway
of human ICC cells. (A,B) Various signaling pathways were detected by qRT-PCR
technology in HuCCT1 (A) and QBC939 (B). (C) Western blotting assays of ERK1/2,
p-ERK, P38 and p-P38 protein expression in HuCCT1 and QBC939 cells treated with
Hinokitiol. Data was analyzed using unpaired t test. N.S., no
significance, *p
Human ICC is a malignant cancer with complex pathology and multiple targets. Combined drug therapy is becoming a trend in tumor treatment [25]. Cyclin-dependent kinase 4 and 6 (CDK4/6) is a promising target for ICC treatment, and Palbociclib, a CDK4/6 inhibitor, has been reported to synergize with pan-mTOR inhibitors to impair ICC growth [26]. As shown in Fig. 6A,B, combined Hinokitiol and Palbociclib treatment showed a much stronger anti-tumor effect than the single drug treatment in clone formation assays (Fig. 6A) and tumor sphere formation analyses (Fig. 6B). Overall, our results suggest that Hinokitiol combined with the CDK4/6 inhibitor Palbociclib could significantly inhibit the clone formation capacity and tumor sphere formation of human ICC cells.
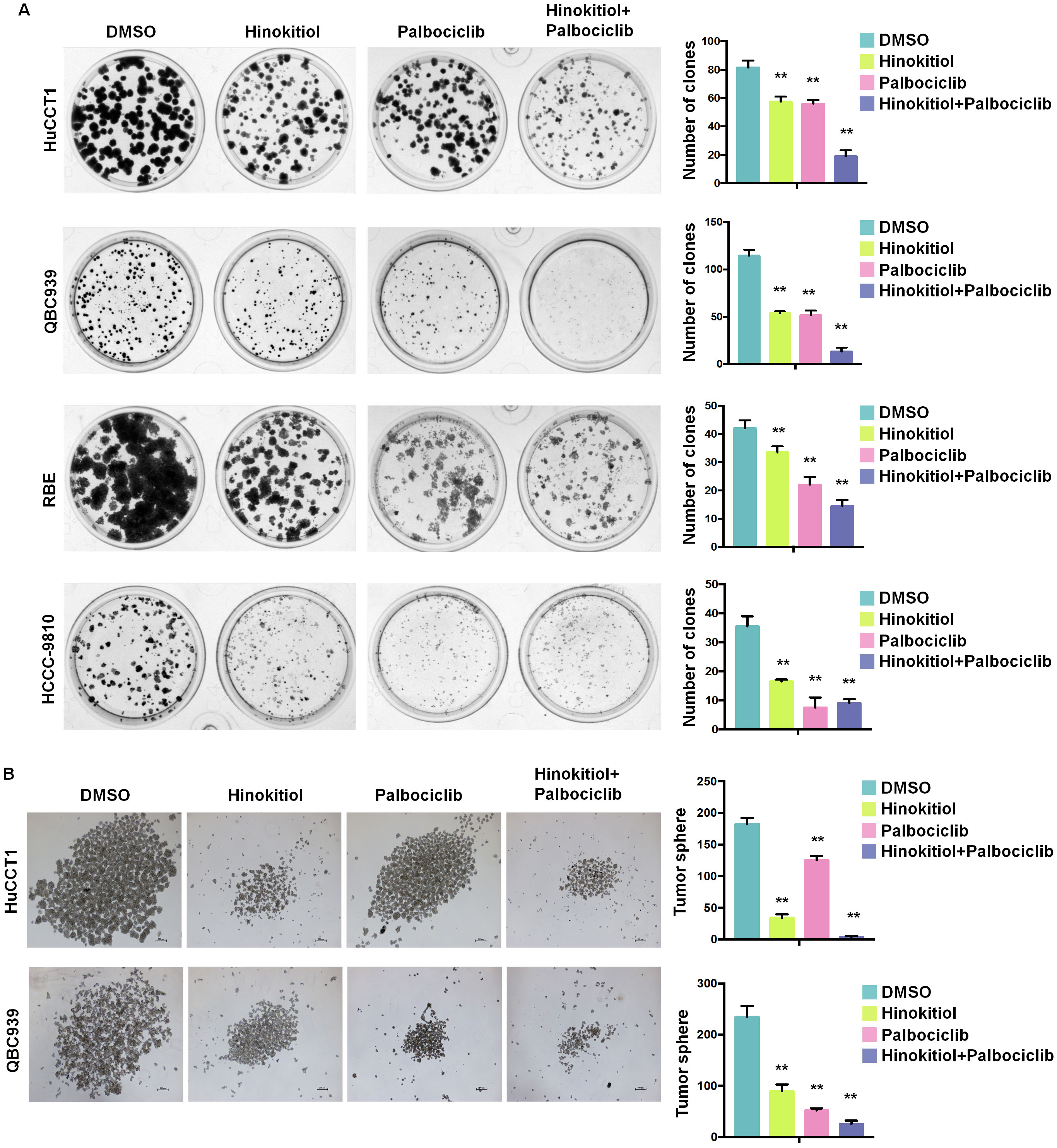 Fig. 6.
Fig. 6.Hinokitiol combined with Palbociclib significantly inhibited
tumor sphere formation of human ICC cells. (A) The effects of Hinokitiol,
Palbociclib, and combined group treatments were evaluated through clone formation
assays in ICC cells. (B) The effects of Hinokitiol alone, Palbociclib alone, and
Hinokitiol + Palbociclib treatments were evaluated by tumor sphere formation
assays in ICC cells. Data was analyzed using unpaired t test. N.S., no
significance, *p
3D tumor organoids derived from human primary tumor tissues and mouse tumor models are novel preclinical model systems that facilitate drug discovery and screening. The ICC mouse model was constructed by hydrodynamic transfection method [27, 28] (Fig. 7A) and was verified by the immunohistochemical staining of CK19 (Fig. 7B). Thereafter, mouse primary ICC cells were purified from the induced mouse ICC tumor tissues and then cultured the mouse ICC organoids in three-dimensional (3D) medium, treated with Hinokitiol, Palbociclib, or combined drug, respectively (Fig. 7C,D). These results indicated that Hinokitiol exerted anti-tumor effects in mouse ICC organoids in vitro. In addition, Hinokitiol combined with Palbociclib significantly inhibited the formation of mouse ICC organoids.
 Fig. 7.
Fig. 7.Hinokitiol combined with Palbociclib significantly inhibited the
formation of tumor organoids from mouse ICC models. (A) The pattern of separation
of tumor organoids in mice ICC. (B) Representative IHC staining of CK19 in ICC
tissues. (C) The formation of mouse ICC tumor organoids was observed after 7
days. Scale bar is 200
In the past 30 years, the incidence and mortality of intrahepatic cholangiocarcinoma have increased worldwide; however, only a few early stage patients can benefit from surgical resection, and most patients diagnosed at an advanced stage have to be treated with standard chemotherapy of platinum or cisplatin combined with gemcitabine; however, the combination treatment of these advanced state patients results in a poor prognosis, with only a median overall survival of 11 months [29]. Therefore, new treatments with better outcomes are urgently required. Current studies have found that low-level reactive oxygen can be an effective mitogen, which is necessary for many biological processes, such as cell survival, proliferation, angiogenesis, and metastasis [9]. Excessive reactive oxygen species metabolically produced by redox transcription factors will affect the therapeutic effect of tumor drugs and lead to drug resistance [30]. Therefore, we utilized a redox library to screen the small molecule compounds to inhibit this refractory malignancy.
In this study, eight small-molecule compounds were firstly selected as candidate
anti-tumor drugs by a preliminary screening stage. Among them,
ICC patients often exhibit high drug resistance to Palbociclib via various mechanisms, leading to an increasing proportion of CSCs and EMT [44]. In this study, we found that Hinokitiol has a better inhibitory effect when combined with Palbociclib treatment, which may play a significant role in the clinical application of ICC treatment.
We highly appreciate that ICC cells from different patients may possess different molecular mutations, such as FGFR, IDH, and BAP. The commonly used ICC cell lines may not be a good model to reflect the response of drugs in ICC patients. In our preliminary experiments, we designed an experiment to culture organoids from human ICC patients using the method of Hidetsugu Saito group [45], but in our hands, we could not successfully culture the human ICC organoids, due to the difficulty of human ICC organoid culture and the scarcity of human ICC fresh samples. Instead, we have successfully cultured the mouse ICC organoids from hydrodynamic transfection-induced mouse ICC tissues, we will consider mutating the mouse ICC organoids to screen the drugs in the future. Actually, Segatto et al. [46] reported that FGFR2 fusion proteins drive oncogenic transformation of liver organoids from mouse ICC tissues.
In conclusion, a natural small molecule compound, Hinokitiol, was found in a redox library that could effectively inhibit the growth of ICC cells. Our results showed that Hinokitiol can effectively inhibit tumor sphere proliferation in vitro by downregulating the expression of the ERK and P38 pathways. Moreover, its combination with the clinical drug, Palbociclib, showed significant anti-tumor effects in human ICC cells and mouse ICC organoids. Therefore, the study of Hinokitiol and Palbociclib provides a promising application for the treatment of patients with ICC.
ICC, Intrahepatic cholangiocarcinoma; CSC, cancer stem cell; qRT-PCR,
quantitative reverse-transcription polymerase chain reaction; HCC, hepatocellular
carcinoma; Nrf2, nuclear factor-erythroid 2 related factor 2; APE-1/Ref-1,
Apurinic-apyrimidinic endonuclease 1/redox factor 1; MAPK, Mitogen-activated
protein kinase; EMT, epithelial-mesenchymal transition; JNK, c-Jun N-terminal
kinase; TGF-
XLY, WY, and XTP conceived and designed the experiments; PB, CG, HY, and HC performed the experiments and analyzed the data; LW, YZ, BZ, QZ, and ZF analyzed the data.
The institutional review board of the Beijing University of Technology approved animal experiments, code 20200311.
We would like to thank Editage (www.editage.cn) for English language editing.
The research was supported by grants from National Natural Science Foundation of China (81772617, 82173183), Great Wall Scholar Project (CIT&TCD20190311).
The authors declare no conflict of interest.